


Dr Souhail Chennaoui, Chercheur INRA CRRA Errachidia
Dans les zones de culture du colza marquées par un stress hydrique accru, il est nécessaire de disposer de cultivars nains et à feuilles lobées étroites afin de réduire la transpiration dans un système de production intensif. Récemment, une lignée mutante naine ‘H2M-1’ et un mutant à feuilles lobées étroites ‘H2M-2’ ont été développés par mutagénèse à l’aide des agents mutagènes chimique (méthane sulfonate d’éthyle, EMS) et physique (rayons gamma) (Channaoui et al., 2019). Pour exploiter correctement ces caractères mutés dans un programme de sélection efficace, il faut comprendre leur mode d’hérédité. D’une part, il existe des résultats contradictoires concernant l’hérédité du nanisme dans les lignées mutantes de colza développées dans des travaux antérieurs (Wang et al., 2016; Xiang et al., 2016). D’autre part, et à notre connaissance, il y a peu d’informations sur l’hérédité et le contrôle génétique de la taille des feuilles (Jian et al., 2017). Par conséquent, l’objectif de cette étude est d’examiner l’hérédité des caractères mutés liés au nanisme et aux feuilles à lobes étroits.
Méthodologie de travail
Des plantes de la variété de type sauvage ‘INRA-CZH2’ (P1) ont été croisées réciproquement en 2015/2016 avec les plantes mutantes ‘H2M-1’ et ‘H2M-2’ (P2) pour obtenir la génération F1. En 2016/2017, les plantes F1 issues de croisements réciproques ont été autofécondés pour obtenir des graines F2 et également rétrocroisées avec les deux parents pour obtenir des graines BC1F1 (F1 × P1 et F1 × P2). Les croisements réciproques originaux ont été répétés pour obtenir des graines F1 dans le même environnement que les graines F2 et BC1F1. Les graines résultantes de P1, P2, F1, F1r, F2, F2r, BC1F1 ont été plantées selon un dispositif complètement aléatoire avec trois répétitions, à la station expérimentale de Sidi Allal Tazi en 2017/2018. Des pratiques culturales conventionnelles et uniformes ont été suivies pour éviter les variations de terrain et les effets environnementaux.
 et feuille lobée étroite du mutant H2M-2 (b). Plante de INRA-CZH2 (c) et plante de H2M-2 (d) au stade de huit feuilles.")
Figure 1. Feuille du variété sauvage INRA-CZH2 (a) et feuille lobée étroite du mutant H2M-2 (b). Plante de INRA-CZH2 (c) et plante de H2M-2 (d) au stade de huit feuilles.
Sur les populations établies, des caractéristiques particulières distinguant les mutants de la variété témoin de type sauvage ont été étudiées, à savoir la hauteur des plantes pour H2M-1 (92,8 ± 4,64 cm) et la forme des feuilles pour H2M-2 (feuille lobée de forme réduite) (Figure 1b, 1d). Le type sauvage a présenté une hauteur de plante moyenne de 161 ± 8,40 cm et une forme de feuille normale (feuilles larges lobées) (Figure 1a, 1c). Pour les populations P1, P2 et F1, dix plantes génétiquement homogènes par population ont été utilisées de manière aléatoire pour toutes les observations et mesures, tandis que pour les populations F2 et BC1F1, le nombre de plantes par population était beaucoup plus élevé, mais variable en raison de la disponibilité de ces plantes comme indiqué dans les tableaux 1 et 2.

Tableau 1. Distribution de la hauteur des plantes et le test du khi-deux pour les ratios de ségrégation dans les populations F1, F2 et BC1F1 résultant de croisements réciproques entre la lignée de type sauvage ‘INRA-CZH2’ et le mutant ‘H2M-1’.

Tableau 2. Distribution de la forme des feuilles et le test du khi-deux pour les ratios de ségrégation dans les populations F1, F2 et BC1F1 résultant de croisements réciproques entre la lignée de type sauvage ‘INRA-CZH2’ et le mutant ‘H2M-2’.
Le test Khi-carré (χ2) a été utilisé pour évaluer les ratios de ségrégation. Ce test permet de mesurer l’écart entre les effectifs observés et théoriques par la formule suivante :
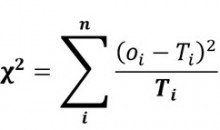
Où,
Oi : effectif observé pour le phénotype i.
Ti : effectif théorique pour le phénotype i.
n : nombre de phénotypes.
Les moyennes réciproques de F1 et F2 ont été comparées par des tests t indépendants afin de trancher sur la présence ou l’absence de l’effet maternel et cytoplasmique sur l’expression et la transmission des caractères mutés.
Résultats
Hauteur des plantes : croisement entre le type sauvage ‘INRA-CZH2’ et la lignée mutante ‘H2M-1’.
 et feuille lobée étroite du mutant H2M-2 (b). Plante de INRA-CZH2 (c) et plante de H2M-2 (d) au stade de huit feuilles.")
Figure 1. Feuille du variété sauvage INRA-CZH2 (a) et feuille lobée étroite du mutant H2M-2 (b). Plante de INRA-CZH2 (c) et plante de H2M-2 (d) au stade de huit feuilles.
Les plantes de la variété INRA-CZH2 sont d’une taille moyenne de 161 ± 8,4 cm, variant de 141 à 180 cm, tandis que celles du mutant H2M-1 font une taille moyenne de 92,8 ± 4,64 cm qui oscille entre 80 et 100 cm (Figure 2). La hauteur moyenne des plantes F1 issues du croisement INRA-CZH2 × H2M-1 est de 157,92 ± 10,1 cm et fluctue de 145 à 175 cm. Cela indique que le caractère sauvage (hauteur normale) est dominant sur le caractère muté (nanisme). Autrement dit, la mutation obtenue est contrôlée par un ou des gènes récessifs. Par ailleurs, les plantes F1 issues du croisement réciproque ont enregistré une taille moyenne de 156,92 ± 9,12 cm, déduite des valeurs allant de 145 à 170 cm (Figure 2). La différence entre les hauteurs moyennes des plantes F1 des deux croisements réciproques n’étant pas significative (t = 0,801, p > 0,05), ce qui indique l’absence d’un effet maternel. La hauteur moyenne des plantes F2 du croisement INRA-CZH2 × H2M-1 est de 146,47 cm et varie de 82 à 180 cm. Cette moyenne n’étant pas significativement différente (t = 0,253, p > 0,05) de la taille moyenne des plantes F2 du croisement réciproque qui est de 146,14 cm et fluctuant entre 83 et 175 cm (Figure 2). L’analyse des générations F2 des deux croisements réciproques confirme ainsi l’absence de l’effet cytoplasmique.
Dans la population F2 regroupée, les plantes ont été divisées en trois classes distinctes pour la hauteur des plantes. La première classe est constituée de plantes ayant une hauteur similaire au parent H2M-1, allant de 80 à 100 cm, la deuxième comprend des plantes de hauteur intermédiaire allant de 104 à 142 cm et la troisième classe est constituée de plantes ayant une hauteur entre 146 et 178 cm similaire au parent INRA-CZH2 (Figure 2). De même, dans chacune des deux populations F2 réciproques, les plantes étaient réparties en trois classes. En effet, le nombre d’individus de la population F2 issue du croisement INRA-CZH2 × H2M-1 dans la classe [80-100 cm] représente environ 1/16, ceux de la classe [104-142 cm] en forment 3/16, alors qu’ils constituent 12/16 dans la classe [146-178 cm]. De même, les individus de la population F2 réciproque suivent la même distribution, avec des proportions d’environ 1/16 pour la première classe [80-100 cm], 3/16 pour [104-142 cm], et 12/16 pour la classe [146-178 cm] (Tableau 1).
Sur la base de ces observations, il a été supposé que deux gènes récessifs à épistasie dominante, contrôlent l’hérédité de la hauteur des plantes de la lignée mutante en question. Les ratios F2 attendus, sur la base de cette hypothèse, seraient de 12:3:1. Le test Khi-carré d’ajustement des ratios observés aux ratios théoriques n’est pas significatif pour les deux populations F2 issues des croisements réciproques (F2 : χ2 = 0,839, p = 0,36 ; F2 réciproque : χ2 = 3,15, p = 0,08) et pour leur agrégation (χ2 = 1,742, p = 0,19), ce qui appuiel’hypothèse de l’épistasie dominante.
L’hypothèse de l’implication de deux gènes récessifs à épistasie dominante est également confirmée par l’analyse de la hauteur des plantes issues du backcross BC1F1 (F1 × P2), pour lesquelles une distribution tri-modale a été obtenue avec des classes distinctes : 80-100 cm, 104-142 cm et 146-178 cm (Tableau 1), et où la ségrégation a suivi les ratios 2:1:1 (χ2 = 1,2, p = 0,27).). Cette hypothèse est de même fortement appuyée par l’examen des plantes issues du backcross BC1F1 (F1 × P1). En effet, 20 plantes sur les 25 analysées se trouvent dans la classe parentale (INRA-CZH2) et cinq plantes ont une hauteur intermédiaire entre 125 et 140 cm (Tableau 1). Cependant, cette ségrégation observée s’ajustait statistiquement aux ratios attendus 0:0:1 (χ2 = 1,00, p = 0,32), confirmant ainsi l’hérédité digénique récessive. L’observation de quelques plantes (cinq plantes) de hauteur intermédiaire dans cette population (Tableau 1) pourrait être due à un effet environnemental et/ou à une erreur expérimentale.
Forme de la feuille : croisement entre le type sauvage ‘INRA-CZH2’ et la lignée mutante ‘H2M-2’.
Les plantes F1 issues du croisement entre INRA-CZH2 et H2M-2 avaient toutes une forme de feuille normale (Tableau 2). Cela montre que la mutation en question « feuille lobée réduite » est déterminée par un ou plusieurs gènes récessifs. Les mêmes résultats ont été trouvés pour le croisement réciproque H2M-2 × INRA-CZH2 (Tableau 2), indiquant l’absence d’effets maternels. En outre, les plantes des deux populations F2 réciproques ont été réparties en plantes à forme de feuille normale et plantes à feuille lobée réduite (Tableau 2). Les proportions respectives observées correspondent parfaitement au rapport attendu de 3:1 (χ2 = 0,23, p = 0,63 ; χ2 = 1,36, p = 0,24), ce qui indique qu’un seul gène est impliqué dans la détermination de la forme des feuilles, avec la dominance de la forme normale des feuilles. Les croisements réciproques ont donné les mêmes résultats (Tableau 2), indiquant l’absence d’implication des facteurs cytoplasmiques dans la détermination de la forme des feuilles. De même, la population F2 regroupée a subi une ségrégation selon le même rapport de 3:1 (χ2 = 0,118, p = 0,73). Les populations rétrocroisées ont bien confirmé l’hérédité simple du caractère muté, due à un seul gène récessif. En effet, la génération backcross fournie par le croisement F1 × INRA-CZH2 est formée en totalité de plantes à feuilles de forme normale, alors que les individus de la génération backcross résultant du croisement F1 × H2M-2 sont à moitié à feuilles de forme normale et à moitié à feuilles de forme modifiée, selon les ratios 1:1 (χ2 = 0,5, p = 0,48) (Tableau 2).
Conclusion et recommandations
Dans ce travail, nous avons étudié l’hérédité des caractères mutés chez un nouveau mutant H2M-1 de type nain et un nouveau mutant H2M-2 de type feuille lobée étroite. Les résultats ont montré que deux gènes récessifs avec une épistasie dominante contrôlent l’hérédité du nanisme, tandis qu’un seul gène est impliqué dans la détermination de la forme des feuilles, avec une dominance de la forme normale du type sauvage. Ces résultats doivent être confirmés et soutenus par des études génomiques et moléculaires. Les lignées mutantes ‘H2M-1’ et ‘H2M-2’ obtenues et caractérisées peuvent être utilisées comme germplasmes utiles dans le programme de sélection du colza.
Références
Channaoui, S., Labhilili, M., Mouhib, M., Mazouz, H., El Fechtali, M., & Nabloussi, A. (2019b). Development and evaluation of diverse promising rapeseed (Brassica napus L.) mutants using physical and chemical mutagens. Oilseeds and fats, Crops and Lipids, 26. 35.
Jian, H., Yang, B., Zhang, A., Zhang, L., Xu, X., Li, J., & Liu, L. (2017). Screening of candidate leaf morphology genes by integration of QTL mapping and RNA sequencing technologies in oilseed rape (Brassica napus L.). PloS one, 12(1).
Wang, Y., He, J., Yang, L., Wang, Y., Chen, W., Wan, S., … & Guan, R. (2016). Fine mapping of a major locus controlling plant height using a high-density single-nucleotide polymorphism map in Brassica napus. Theoretical and Applied Genetics, 129(8), 1479-1491.
Xiang, Y., Tong, C., Yu, S., Zhang, T., Zhao, J., Lei, S., … & Liu, S. (2016). Genetic Segregation Analysis of a Rapeseed Dwarf Mutant. Pakistan Journal of Botany. 48(4). 1629-1635.
——————————————
1 INRA, Centre Régional de la Recherche Agronomique de Meknès
2 INRA, Centre Régional de la Recherche Agronomique d’Errachidia